Дипломная работа: Розмірно-віковий та статевий склад популяціі ляща
Вступ
В останні роки в водосховищах Дніпровського
каскаду спостерігається зниження запасів основних промислових видів риб, таких
як лящ, плітка, судак, короп. Це викликано значним антропогенним навантаженням
на популяції цінних промислових видів риб: Промислове та любительське
рибальства, процвітаюче браконьєрство, забруднення промисловими стоками різного
походження. В результаті замулення таких водосховищ як Каховське, поступово
витрачаються значущі нерестові площі, що знижує ефективність природного
відтворення водних живих ресурсів.
Каховське водосховище – одне з найбільших на
Дніпрі, що замикає каскад і розташовано на самому півдні річки. При проектуванні
Каховського водосховища площею 215 тис. га відповідно опрацювань інституту
гідробіології АН УССР планувалось щорічно отримувати 10 тис. тон риби, в тому
числі до 80% цінних видів риб – судака, ляща, сазана, сома, щуки.
Після перекриття Дніпра в 1955 році Каховською
греблею в період заповнення водосховища і залиття великих площ заплав утворились
сприятливі умови для нересту, що забезпечило збільшення їх вилову до обсягів
планових показників в 1972 році. Частка в уловах цінних риб досягла 50%.
Тому раціональне використання біоресурсів
Каховського водосховища неможливо без знання основних характеристик та
особливостей екосистем, для чого необхідно постійне вивчення іхтіофауни
водоймища. На підставі даних, отриманих при зборі й обробці іхтіологічного
матеріалу можна судити про запаси промислових видів риб, умовах відтворення риб
на акваторії Каховського водоймища, давати оцінку біологічного стану нерестових
популяцій, визначити строки весняно-літньої заборони. На підставі цього
здійснювати правильні корегування промислу. Особливо це стосується ляща, як
одного з вагомих об’єктів рибного промислу.
Метою даної роботи було дослідити
розмірно-віковий та статевого склад популяції ляща як об’єкта промислового
рибальства в Каховському водосховищі.
Перед нами стояли такі завдання:
- Визначити
розмірний склад популяції ляща;
- Визначити віковий
склад популяції ляща Каховського водосховища;
- Дослідити
статевий склад популяції та плодючість ляща Каховського водосховища;
Ця робота
актуальна у зв’язку з тим, що раціональне використання біоресурсів Каховського
водосховища неможливе без знання основних характеристик та особливостей
екосистем, для чого необхідно постійне вивчення іхтіофауни водоймища.
На підставі
даних, отриманих при зборі й обробці іхтіологічного матеріалу можна судити про
запаси промислових видів риб, умовах відтворення риб на акваторії Каховського
водоймища.
За результатами
дослідження можуть бути розроблені біологічні обґрунтування використання
промислової іхтіофауні Каховського водосховища.
1. Огляд наукової літератури
1.1 Характеристика ляща
Лящ (Abramis brama) відноситься до роду
абраміс (Abramis), ряду коропообразні (Cyprinipormes).
Тіло ляща помірно видовжене, високе, досить
стиснуте з боків. У бічному профілі, особливо у дорослих особин, помітно різке
підвищення тіла одразу за потилицею. У самців і самок забарвлення однакове –
переважно сріблясто-сіре на боках з потемнінням на спині й посвітлінням на
череві. Плавці темнуваті.
Рогівка ока сіро-золотиста. Кольорові відтінки
тіла змінюються з віком: від світло-сірого з сріблястістю у молоді до
бурувато-темного з золотисто-жовтуватим відливом у дорослих особин. При цьому з
віком риб у них проступає рожевуватий колір на череві і під горлом, особливо
навесні. Плавці із сірих з чорнуватими кінцями стають цілком темними. Рогівка
ока із золотистої стає коричнюватою [1,2].
Прісноводна, іноді солонуватоводна, річкова і
озерна, напівпрохідна і туводна, придонна, мало реофільна фітофільна зграйна
мирна риба. Живе у річкових басейнах, переважно в середніх і нижніх частинах
рік з повільною течією, не поширюючись у невеликі ріки гірського типу з швидкою
течією і холодною водою. Тяжіє до прісних, повільно текучих вод, зустрічається
в опріснених ділянках морів (Балтійського, Чорного і Азовського). Далеко в море
не виходить. Солоність вод, в яких зустрічається лящ, становить за хлором не
вище 2,5–3 г./л, або до 5–6 ‰[2].
Забарвлення змінюється залежно від якості води й
кольору ґрунту. У торф'яних озерах і ставках лящ темніший, майже бурого
кольору, у ріках і водоймах з піщаним дном – світліший, з золотистим відливом
[3,4]. На середній течії Дніпра відмічається варіація кольору тіла ляща залежно
від того, до якої групи він належить: лящ-вербовик має жовтуватий колір,
лящ-дубовик – темно-сріблястий, лящ-плоскирник – світло-сріблястий. У ляща, що
нереститься ранньою весною, забарвлення світліше, ніж у ляща, що нереститься
пізньою весною. Статевий диморфізм виражений слабо[5].
Відомий у Європі на схід від Піренеїв і на північ
від Альп, річкових басейнах і опріснених ділянках Північного, Балтійського,
Білого, Егейського, Чорного, Азовського, Каспійського морів, Ірландії, Англії,
пониззі Рони. У Швейцарії зустрічається тільки в басейні Рейну, у Фінляндії
всюди в басейні Балтійського моря, у Фінській затоці всюди, але переважно у
східній опрісненій частині. Звичайний у прісних водах басейну Білого моря – в
Топозері, Керетьозері, Сегозері, Чарандському озері, у Північній Двіні від
Кубенського озера (у великій кількості) до Архангельська, в ріках Кулой, Мезень
і Печора. На півдні відомий у пониззі Струми, Пенея, пониззі Ріоні (оз.
Палеостомі). Ца схід від Печори, зокрема в Сибіру, невідомий. Акліматизований у
системі верхів'я р. Ісеть на східному схилі Уралу і в окремих водоймах Сибіру,
в Убинському озері (басейн Обі), в Іртиші (біля Тобольська) і в оз. Балхаш [6].
1.2 Екологія ляща
У річкових басейнах лящ тяжіє до заток, бічних
проток, проточних озер. Характерний лящ і для великих озер. Типово лящеві озера
– евтрофні водойми з добре вираженою літораллю. В дельті Дунаю лящ
представлений у різних ділянках, але в більшій відносній кількості у великих
затоках («кутах») (46,12% загальної кількості риб у контрольних виловах), в
трохи меншій кількості – на мілинах передгирлових ділянок дельти (41,3%) і в
значно меншій кількості – в протоках-гирлах (6,8%) і заплавах (5,7%). Зовсім
відсутній у невеликих протоках і затоках [7, 8, 9, 10, 11].
Лящ досить легко пристосовується до умов
зарегульованого річкового стоку і знаходить сприятливіші умови для життя у
річкових водоймищах. Добре приживається й у неспускних ставах і навіть у
спускних (до осені) [12]. Типові для ляща ріки відрізняються твердо-піщанистим,
злегка замуленим або й глинистим дном. Уникає лящ місць з рівним піщаним дном
або дуже замуленим і зарослим рослинністю. Якщо і буває в них, то спорадично,
при переходах з місця на місце. У характерних для ляща ріках дно утворює ями на
поворотах, під ярами, має уступи, що захищає ляща від течії і різної небезпеки.
Звичайним місцем перебування ляща, особливо
старшого віку, є глибокі заводі під обривистим берегом з ущільненим
мулувато-піщаним або глинистим дном з уступами, поглибленнями, глинистими
грудками, корчами, завалами. Зрідка знаходиться біля кам'янистих або піщаних
мілин. Досить охоче тримається в ямах біля мостів, між палями. У невеликих
ріках з глинистим або чорноземним берегом лящ, особливо його молодь,
зустрічається у прибережній смузі, де течія вимиває з берега кормові організми.
У більшій частині озер і в ставках лящ тримається відносно глибоких, дещо
замулених, більш-менш віддалених від берегів і заростей ділянок, іноді біля
кромки заростей на чистому плесі. Молодші особини ляща довжиною до 10 см
частіше тримаються прибережної смуги з негустими заростями, старші особини
більших розмірів відходять від цих місць і тримаються глибших ділянок водойми
(ріки, озера тощо). У пониззі Дніпра з літній період молодь ляща поширена по
всій заплаві.
Лящ – дуже обережна, ляклива риба. Не дуже
рухливий, він переважно знаходиться біля дна в тихих і відносно глибоких
місцях. Періодично виходить на мілини, здебільшого в тихі й похмурі дні, іноді
у спеку, особливо перед грозою, також ночами. Тяжіє до чистої свіжої води,
особливо тої, що тече з лісових джерел. Протягом доби лящ активний у світлий
період, переважно вранці до 11 години, частково й уночі, зокрема в середині
літа.
Значних міграцій лящ не робить, хоча міграційна
поведінка властива всім популяціям ляща. Вираженість міграцій у ляща
визначається належністю його популяцій до певної екоформи: напівпрохідної або
туводної. Туводній (в тому числі й озерній) формі властивий осілий спосіб
життя, напівпрохідній – мігруючий. Всі життєві цикли у туводної форми проходять
в певній ділянці ріки, у напівпрохідної форми місця нагулу, зимівлі й нересту
більш-менш віддалені одне від одного. Існування двох форм відмічено для різних
рік, у першу чергу для тих, які мають досить велику естуарну частину (Дністер,
Південний Буг, Дніпро, Дон, Волга тощо). Ці форми не розділені географічно, і
напівпрохідна форма часто досягає місцезнаходження туводної.
В результаті зарегулювання річкового стоку
зникають необхідні умови для напівпрохідної форми і виникають умови для
існування туводної. Зокрема, в Дністровському лимані при зарегулюванні Дністра
відмічена поступова заміна напівпрохідної форми туводною [13, 14, 15]. В районі
Каховського водоймища до зарегулювання пониззя Дніпра стадо ляща було
неоднорідне. Існував напівпрохідний лящ, що мігрував від гирла до Запоріжжя.
Відмічалось стадо і туводного ляща на запорізькій ділянці й аналогічне стадо в
дельті Дніпра. Після утворення Каховського водоймища в ньому сформувалось
велике і біологічно різнорідне стадо, складене напівпрохідною і туводною
формами, причому в подальші роки існування водоймища остання стала домінуючою
[15, 16, 18,].
Таким чином, при характеристиці міграцій ляща
мова може йти в основному лише про напівпрохідну форму виду. В ріках, зокрема в
Дніпрі, у ляща відмічаються два чітко виражені ходи з лиманів: у березні-квітні
й у вересні-грудні. В ріках плідники ляща одразу після нересту у заплаві з вище
розташованих ділянок більш-менш активно мігрують униз за течією на нагул в
лиманну частину або в опріснену ділянку моря. Ця міграція починається на
початку травня, стає масовою наприкінці його – в перших числах червня і
закінчується в першій половині липня. Міграція, або скочування, має більше
індивідуальний характер, ніж косячний, оскільки плідники закінчують нерест
неодночасно і, не затримуючись на нерестовищі, йдуть вниз за течією [19, 20].
Із молоді ляща лише невелику частину личинок
одразу після викльову з ікри течія виносить із заплави ріки і зносить вниз.
Звичайно молодь спочатку залишається в місцях нересту плідників, тримаючись
мілководдя з помірно розвинутою рослинністю потім переходить в глибші ділянки
ріки і під кінець літа током води, тобто способом пасивної міграції, частково
активно, зноситься в естуарну частину річкового басейну або в опріснену річковими
водами ділянку моря. Проте частина мальків, не встигнувши скотитись,
залишається у дельті ріки і вище розташованих її ділянках на зимівлю.
Скочування молоді триває з початку липня до кінця серпня, іноді до кінця осені.
У Дніпровсько-Бузькому лимані молодь нагулюється у прибережній зоні всюди і при
досить різній солоності води. Нагул триває до настання статевого визрівання з
перервами на зимівлі, які проходять у передгирлових ділянках.
Зимує лящ у глибоких ділянках річкових гирл та
прилягаючих естуарних ділянках. У Таганрозькій затоці лящ зимував у
південно-західній частині і в гирлах Дону, в Єйському лимані, зрідка біля кіс
Білосарайської і Кривої та біля Таганрогу. Взимку при дуже низькій температурі
води лящ залягає на ями, зовсім втрачає активність, вкривається досить товстим
шаром слизу. У теплу зиму зберігає певну рухливість, особливо у відлигу і
навіть потрапляє на гачок любителів підльодного лову, що свідчить про потребу
його у їжі. За зиму лящ витрачає певний запас енергетичних ресурсів. Зокрема,
кожен грам його живої маси за час зимівлі втрачає від 0,13 до 0,23 г., залежно
від суворості зими [21].
Післязимове поновлення активності ляща
відмічається з першими ознаками ранньовесняного потепління ще під кригою. Вже з
кінця лютого, особливо на півдні, лящ виходить із зимувальних ям на
мілководніші ділянки, живиться і, збиваючись у косяки, починає переміщуватись у
розташовані вище ділянки рік. У пониззі Дніпра до спорудження Каховської ГЕС
хід ляща з лиману в ріку починався майже завжди незабаром після скресання
криги, проте до середини квітня тривав слабко, з перервами.
Співвідношення статей залежить від віку риб: у
молодших вікових групах чисельно переважають самці, у старших – самки. Так, у
пониззі Дніпра в 1976 р. з переходом по вікових групах від 5 до 11 років
відмічалось зменшення відносної кількості самців від 58,9 до 0% і збільшення
відносної кількості самок від 41,1 до 100% [15, 22].
Абсолютна плодючість із зростанням розмірів і
віку самок збільшується. Приблизно підраховано, що із зростанням довжини тіла
на 1 см кількість ооцитів збільшується на 10,2 тис. ікринок, а із
зростанням маси тіла на 100 г. – на 9,5 тис. У дев'ятирічних самок плодючість
вп'ятеро більша, ніж у трирічних. Кореляція між довжиною тіла і абсолютною
плодючістю досягала 74%. Із збільшенням маси тіла від 400–600 до 1400–1600 г.
абсолютна плодючість збільшувалась від 46,1 до 236 тис. ікринок.
Місця нересту ляща звичайно знаходяться вище за
течією від місць зимівлі й основного нагулу, у мілководних ділянках прибережних
водойм, у лиманах, протоках, затоках, залитих ділянках заплавної тераси,
заплавних озерах. Тут грунт дна звичайно твердий, незамулений (піщаний), з
плямами помірно розвинутої рослинності. Зарегулювання річкового стоку
позначається на місцях нересту. Так, у пониззі Дністра в зарегульованих умовах
і значного (штучно викликаного) коливання рівня води лящ широко освоїв замість
заплави нові нерестові ділянки і нерестовища на більшій глибині в Дніпровському
лимані, в його найопрісненіших ділянках. При цьому частина стада ляща
(напівпрохідний) нереститься у дельті Дністра, а друга частина (туводний) – на
заплаві [23].
Після зарегулювання стоку Дніпра у водоймищах
його середньої течії субстратом для нересту ляща часто є залишки торішньої
рослинності, затоплений чагарник, молоді паростки очерету, також обвітрене
коріння верби тощо. У Дніпровсько-Бузькому лимані для ляща характерне
відкладання ікри переважно на дно, на коріння вищої водної рослинності [16].
Строки нересту залежать від гідрометеорологічних
умов. Час нересту ляща зонально пристосований, що зумовлено початком весняного
прогріву водної товщі. У південних районах нерест відбувається раніше – з
третьої декади квітня до початку червня, у північних – пізніше, у
травні-червні.
За сприятливих умов нерест проходить бурхливо.
Нерестуючі. групи плавають біля нерестовищ, окреслюючи коло і поступово
звужуючи його з діаметра 8–4 м до мінімуму. Плідники рухаються дуже
активно, особливо самці, іноді вискакують над водою і з плескотом падають
назад. Бурхливо нереститься лящ і на глибині, хоч при цьому й менше помітно.
Під час нересту лящ інтенсивно треться об субстрат, про що свідчить загублена
на субстраті луска.
При нересті самка в супроводі кількох (зрідка
одного) самців снує серед рослинності. При потужних рухах тіла самки у неї
тонкою майже безперервною цівкою витікає ікра, яку вона розсіює на субстрат, а
самці тут же поливають її молоками. Самка відкладає всю ікру за 1–2 год. В
середньому одна самка відкладає до 200 тис. ікринок. Самці можуть в один сезон
кілька раз брати участь у нересті.
У природних умовах самки ляща відкладають ікру
нещільно, поодиноко на досить широкому просторі (хоча загальна площа місця,
засіяного ікрою, не перевищує 0,01–1 га), в місцях, де є рослинність і глибина
порівняно невелика (до 2 м). Тут, як правило, не буває дефіциту кисню,
ікра знаходиться в верхніх шарах води, де вміст кисню достатній. Крім того,
рослини при фотосинтезі вдень виділяють кисень, збагачуючи воду. Розсіяна
кладка ікринок сприяє вільнішому доступу до них кисню.
У Каховському водоймищі молодь ляща довжиною до
10 см живиться рослинним детритом (42,2% маси їжі) і нижчими
(гіллястовусими) ракоподібними (решта їжі). При досягненні 5–10 см молодь
переходить на живлення придонними формами нижчих ракоподібних (27,5%). в
значній мірі поїдає детрит (до 41,4%), личинок хірономід (до 12,7%), у
невеликій кількості синьозелені водорості тощо (4,8%). У риб завдовжки 10–15 см
харчовий спектр майже не змінюється, але зменшується значення нижчих
ракоподібних (Сореросіа і Сіасіосеrа), зростає роль личинок хірономід та
інших комах, особливо їх імагінальних стадій, а також детриту (до 53,5%).
Молодь, довша від 15 см, живиться також переважно донними та придонними
організмами (13,3%), як домішок у них трапляється пісок (12,7%) [10, 21].
1.3 Фізико-географічна характеристика району досліджень
Дослідження проводились в верхній частині
Каховського водосховища – одного з найбільших штучно створених водних об'єктів
Європи, статус якого, як водойми комплексного призначення, передбачає
інтенсивну рибогосподарську експлуатацію.
На відміну від інших рівнинних водосховищ
України, Каховське водосховище відрізняється відносно невеликою площею
мілководних ділянок складає біля 5% площі водного дзеркала, а також високим
рівнем антропогенного впливу. Наявність промвузлів, водозаборів (в тому числі
великої потужності), нижнє розташування в каскаді водосховищ Дніпра – все це
істотно впливає на екологічну ситуацію у водоймі та значною мірою визначає
структурно-функціональні показники іхтіоценозів.
Каховське водоймище утворилося в результаті
зарегулювання нижньої течії Дніпра греблею ГЕС біля м. Нова Каховка. Площа
водойми становила 215 тис., га, протяжністю 230 км, середня глибина – 8,4 м
(максимальна 38 м). Глибини менше 2 м займають лише 5% її площі, від
2 до 5 – 18% і більше 5 – 77%.
По глибинам та характеру ложа водоймище поділяють
на три ділянки: верхній мілководний та широкий, з глибинами 3–5 м та
шириною, досягаючою в районі затоплення Кінських плавнів 25 км; середній –
від м. Марганець до сіл Балабино, Малі гирла, з глибиною 10–12 м та
шириною 8–13 км, і нижній – найбільш вузький (5–6 км), з глибинами 13–26 м.
В залежності від глибин та характеристик ложа
водоймища існують основні місця розмноження промислових риб. Основні місця
розмноження промислових риб верхньої частини Каховського водоймища знаходяться
в прибережній зоні його заток, мілководних ділянок і вздовж побережжя островів.
Загальна площа найбільш значущих в цьому відношенні заток складає 288 га, мілководних ділянок – 4 728 га та мілководь прибережної лінії островів – близько 380 га [24].
Значення всіх мілководних ділянок вершини
водоймища для риби із року в рік знижується, що обумовлено в основному
забрудненням цих ділянок стічними промислово-побутовими водами м. Запоріжжя,
руйнуванням та розливом берегів, заростанням прибережної зони водною
рослинністю, заболочуванням.
Після спорудження Каховської греблі і утворення
водосховища умови життя даної фауни в нижній течії Дніпра різко зменшилися. В
Каховському водоймищі в 1-й рік його утворення виникли особливо сприятливі
умови для масового розвитку багатьох безхребетних тварин: дном водосховища
стала родюча долина Дніпра з величезною кількістю різноманітної рослинності, що
почала поступово загниватися, даючи цим самим корм і мікроорганізми для водних
тварин.
Відносно невелика глибина (5–0 м) на значних
площах водосховища сприяла перемішуванню води, аерації природних пластів і
утворенню достатньо хорошого гідрохімічного режиму, внаслідок чого досягло
доволі великого численного розвитку багатьох безхребетних тварин, в тому числі
і зі складу фауни лиманно-морського комплексу (каспійського типу) олигохети,
дрейссени, мізіди, гаммаріди, личинки тендипедид, поденок і інших водних комах.
Донна фауна нижньої течії Дніпра від м. Запоріжжя
до м. Нікополя (вище підтопу Каховського водосховища) входить до складу
чотирьох типів біоценозів: псамморефильних, пелореофильних, літореофильних і
фитореофельних; перші два являються пануючими. Біомасу фауни піщаних ґрунтів
склали три групи тварин: олігохети, молюски і личинки тендипедид. Максимальна
чисельність фауни піщаного дна – 2 280 екз/м, біомаса – 16,944 г./м2.
Спорудження Каховської греблі і утворення
водосховища дуже позначилося на зміні якісного складу і численного розвитку
даної фауни. Замість господарюючих псамморефильних біоценозів нижньої течії
Дніпра почали формуватися доволі багаті пеле – (рео) – фільні та фіто – (рео) –
фільні біоценози водосховища.
Спорудження Каховської греблі вносить глибокі
зміни в рибне господарство і рибну промисловість нижнього Дніпра як у
відношенні якісного і кількісного складу промислових уловів, так і в
організаційних рибоводних та риболовних закладах.
Розташована менш ніж у 100 км від гирла
Дніпра, гребля зупиняє доступ до нерестовищ прохідних риб: оселедців, осетра,
білуги і севрюги, а також в значній мірі й напівпрохідних риб: тарані, чехоні,
ляща, судака й рибця (особливо перших двох). В цьому відношенні вплив
Каховської греблі на і якій мірі не може ототожнюватися з впливом греблі
Дніпрогесу, так як остання значно віддалена від Дніпровсько-Бугського лиману,
який є місцем началу напівпрохідних риб, розташована вище основних нерестовищ
перерахованих риб і не спричинила будь-якого помітного впливу на їх запаси.
Каховська гребля обумовлює перетворення значної
частини нижнього.
Дніпра, на протязі 235 км, в величезне
водосховище, ширина якою місцями досягає 20–23 км, а площа водної поверхні
при нормальному підпірному горизонті на відмітці +15,0 становить 222000 га при об'ємі води 16,7 м. В маловодні роки річна витрата води водосховища виражається в
зниженні рівня води з відміткою +15,0 до 10,5, що скорочує площу водного
дзеркала до 150000 га, а об'єм води – до 6,5 км3.
Рибна продуктивність водойм визначається багатьма
факторами; основними з них є умови розмноження риб та стан кормових
ресурсів. Крім форми господарства (включаючи промисел) та флюктуацій, як
зазначив Є.В. Боруцький (1961), одним з основних факторів, який визначає
темп росту та кількість промислових риб, є забезпеченість поживою. Величина
популяцій виду тим більша, чим більші кормові ресурси, які можуть бути
використані видом.
Починаючи з 1958 р. умови для розмноження
риб в Каховському водоймищі різко погіршилося. Скоротилася площа заростей вищої
водної рослинності, в основному закінчилися процеси мінералізації затопленої
лучної рослинності. Правда, рівень води в 1958 р. був надзвичайно великий
(0,75 – 1,0 м вище рівня попередніх років), в результаті чого з'явилися
нові затоплені площі, що сприяло кращому нерестові риб. Але разом з тим різкий
підйом рівня води, що відбувався в період нересту (травень-червень), негативно
відбивався на нормальній інкубації відкладеної ікри.
Каховське водоймище, як і багато інших, з
третього року свого існування характеризується недостатністю нерестових площ
для фітофільних риб та несприятливими умовами нересту через коливання рівня
води. У зв'язку з цим треба було провести ряд меліоративних заходів,
спрямованих на відтворення рибних запасів. Позитивне значення для формування
іхтіофауни водоймища мали роботи Українського інституту рибного господарства.
Найбільшу потенціальну рибопродуктивність
одержано для 1957 р. – 793,4 кг/га. Вона є характерною для удобрюваних ставків.
Це цілком зрозуміло, бо в перші два роки ефект добрива від розкладання
затопленої рослинності та вилугування грунтів у Каховському водоймищі був
надзвичайно великий. У наступні два роки (1958–1959) потенціальна
рибопродуктивність значно зменшилася: у 1958 р. вона становила 559,5
кг/га, а в 1959 р. – 301,3 кг/га. А промисловий лов риби у Каховському
водоймищі почався в 1960 р. [4].
З утворенням Каховського водоймища почали
відбуватися зміни в заростанні водоймищ нижньої течії Дніпра. Заростання Каховського
водоймища відбувалося в два етапи. Перший етап – формування рослинності
водоймища – зайняв період від початку затоплення до утворення водних заростей
водоймища, і другий етап – період розвитку заростей водної рослинності в
утвореному водоймищі.
Для першого етапу – періоду формування характерні
наступні явища: відмирання наземної рослинності в результаті затоплення,
вегетація раніше існуючої водної рослинності в умовах підвищення рівня в
зв'язку з підтопленням, заселення водоймища водною рослинністю.
Відмирання наземної рослинності. Затоплення було
негативним фактором в житті наземних рослин. Протяжність половоддя та висота
полої води в низинах рік призвели до відбору незначної кількості видів, причому
різні види реагували на затоплення неоднаково, що призвело до формування
різноманітних рослинних угрупувань на різноманітних рівнях. При формуванні
водоймища різноманітні рослинні угрупування та окремі види відмирали в різні
терміни. Деревовидна та чагарникова рослинність не переносять тривалого
затоплення, цим і пояснюється безлісся луків. Найбільш стійкі деякі види верб (Salix
alba L., S. Amygdalina L), але і вони при затопленні водоймища відмерли у
перші два роки. Рослинність високих та середніх луків, яка складалася з
короткопойменних рослин, загинула в перший же вегетаційний сезон.
Як показали спостереження в поймі в період
половоддя, підвищення рівня затримує розвиток водної рослинності. Спостереження Д.О. Свиренко
[8] на озері Леніна в період затоплення показали, що водна рослинність відмерла
в перший же рік підтоплення, хоч частина її вегетатувала в перший рік і
загинула тільки на другий рік затоплення.
Спостереження, проведені на ряді побудованих
водоймищ (Магнітогорське, Іваньковське, Шошенське, Рибінське та ін.), в тому
числі і на Каховському, озері Леніна, показали, що заселення мілководних
ділянок водною рослинністю відбувалося дуже швидко. Водна рослинність з'явилася
вже в перше літо після наповнення водоймища, особливо інтенсивно заростали
впадаючі в нього мілководні затоки та балки. Та і на основному плесі водоймища,
в його мілководних ділянках, особливо біля островів, водяна рослинність
з'явилася вже в перший рік. В складі рослинності головну роль грали занурені
рослини і серед них в першу чергу – звичайні для Дніпра види: рдести
(пронизанолистний, гребінчатий, різнолистний, кучерявий, блискучий),
роголистник занурений, уруть колосиста, гречка зеленоводна та інші.
Напівзанурені водні рослини в перший рік
з'явилися на багатьох ділянках водоймища, але в його заростанні вони відіграли
меншу роль. З напівзанурених з'явилися сусак зонтиковидний, очерет озерний,
тростник, кілька видів рогози, стрілолист звичайний та інші [5].
Головним фактором, який визначає розповсюдження
вищої водної рослинності в водоймищах уповільненого стоку, є глибина. На основі
вивчення заростання великого числа озер і водоймищ Г.І. Долгов [4] приймає
максимальну глибину зони заростання в водоймищах в 2 м. Заростання за
межами вказаних кордонів, на глибину 3–4 м, носить випадковий характер,
розповсюджуючись лише клинами та невеликими ділянками.
На основі даних по заростанню Каховського
водоймища встановлено, що головну роль тут зіграли підводін рослини, звичайні
для Дніпра: рдести, роголистники, уруть, підводні форми стрілолиста і сусака, а
також напівзанурені рослини. Рослини з плаваючим листям розвинулися в деяких
місцях як залишки існуючої водної рослинності.
Побудова Каховської греблі різко зменшила витрати
Дніпра і сумарний стік нижче греблі. В середній по водності рік стік нижче
греблі падає в квітні, в травні, в червні, і тільки в серпні зарегульований
стік дорівнює незарегульованому стоку в цім місці. З побудовою греблі різко
зменшилися весняні розливи на Дніпрі нижче греблі.
Як виявили спостереження по впливу половоддя на
розвиток рослинності в заплаві, половоддя чинить затримуючий вплив на хід
вегетації. В один і той же рік в залежності від висоти води на різних
водоймах одні і ті ж види раніше починали вегетацію, в яких рівень і тривалість
половоддя була меншою. Таким чином, зниження половоддя у зв'язку з побудовою
Каховського водоймища зробило благотворну дію на розвиток рослинності.
Збільшення зростання водоймища рослинністю значно погіршує його
рибогосподарське значення [6].
Каховське водоймище, одне з найбільших не лише в
Україні, а і в колишньому СРСР, має велике значення в рибопромисловості країни.
У найближчі роки завдяки цілому ряду рибоводно-меліоративних заходів вилов риби
в ньому повинен досягти до 100 тис. ц.
Проте у робітників рибної промисловості
викликають тривогу випадки загибелі риби (сазана, ляща, судака та ін.), що мали
місце у водоймищі. Так, у 1956 р. загибель риби була помічена на ділянці в
20 км від с. Велика Знаменна до с. Ушколка по східному березі середньої
ділянки водоймища, від с. Гаврилівка до с. Мигове по західному березі та в
інших місцях. Риба гинула від замору – відсутності достатньої кількості
розчиненого у воді кисню.
В серпні 1960 р- в поверхневих шарах
у зв'язку з цвітінням (при інтенсивному фотосинтезу) в штильову погоду
спостерігалося перенасичення води киснем до 200% і більше. В придонних шарах у
зв'язку з вживанням кисню на окислення органічних речовин свіжозалитої землі,
орних угідь і рослинного покрову його вміст знижувався. Так, у серпні в
поверхневих шарах вміст кисню коливався від 5 до 12 мг/л (65–135% насиченості),
а в придонних – від 4 до 7 мг/л (40–75%).
Найбільш сприятливий кисневий режим спостерігався
в верхній ділянці водосховища (на колишніх плавнях). В середній ділянці
водосховища в придонних шарах вміст кисню не перевищував 60–65% насиченості, а
на ділянці біля греблі – 45–50% (в поверхневих шарах коливалося від 80 до 105%
насиченості).
Восени 1956 р. у зв'язку з інтенсивною
вітровою діяльністю, вміст кисню став рівномірним по всій товщі води і майже на
всіх ділянках водоймища досягав 75–85% насиченості [25, 26].
Кормова база риб Каховського водосховища протягом
останніх років характеризувалась достатньо високими показниками.
Якісний склад фітопланктону водойми в літній
період був представлений 58 таксонами, серед яких переважали зелені (29 видів)
і діатомові водорості (18 видів).За чисельністю домінували синьозелені
водорості (основні форми – Microcystis, Anabaena, Aphanizоmenon).
Середньорічна біомаса фітопланктону в літній період може біти оцінена в 9,4 г/м3,
основу якої складали синьо-зелені, діатомові (Melosira granulata, M. іslandica)
та зелені (протококові) водорості. Загальна продукція фітопланктону за
вегетаційний сезон на ділянці, що відводиться для ведення СТРГ, може бути
оцінена 18800 кг/га, потенційний приріст іхтіомаси риб-фітофагів – 376 кг/га,
що відповідає високому рівню кормності.
Загальна біомаса зоопланктону Каховського
водосховища в останні роки відносно стабільна і коливалась в межах 0,9–2,1 г/м3.
Спостерігалось характерне для водойм з закритою вершиною нарощування біомаси
від вершини до пониззя. На всіх ділянках водосховища домінували веслоногі рачки
та велігери дрейссени, складаючи, відповідно 49 та 33% від загальної біомаси.
Серед веслоногих домінуючими були Diaptomus gracilis, Diaptomus graculoides.
Доля гіллястовусих у водоймі була значно менша (13%), найбільше значення вони
мали у вершині водойми (19%). Серед гіллястовусих домінуючими були такі види Bosmina
longirostris, Bosmina coregoni, Chidorus sphaericus. Досить незначною була
доля коловерток у водоймі, становлячи всього 5%, серед них найбільш поширеними
були такі види Asplanhna priodonta, Кeratella guadrata, Кeratella
cochlearis. Середньорічна біомаса зоопланктону становить 1,33 г./м3,
що відповідає середньому рівню кормності. Загальна продукція за вегетаційний
сезон становить 2048 кг/га, що забезпечує потенційний приріст іхтіомаси на
рівні 85,4 кг/га.
Фауна дна верхньої частини Каховського
водосховища характеризується переважанням пело(рео) фільних біоценозів, до
складу яких входять олігохети, поліхети, гаммариди, личинки комах, молюски.
Домінуючою групою організмів серед «м’якого бентосу» ділянки водосховища, що
розглядається, були представники вищих ракоподібних, серед яких домінували
представники родів Pontogammarus, Dikerogammarus, Aselus. Серед комах
основними формами були представники родин родини Chironomidae, Tipulidae.
Інші групи «м’якого» бентосу організмів із-за їх незначної кількості та біомаси
значної ролі для живлення бентофагів не відіграють. Середньовегетаційна
чисельність «м’якого» зообентосу Каховського водосховища становить 795 екз/м2,
біомаса – 2,25 г./м2, що відповідає середньому рівню кормності. Для
кормових молюсків ці показники становили відповідно 176 екз/м2,
біомаса – 8,1 г/м2. Продукція кормового зообентосу становить 210
кг/га, потенційний приріст іхтіомаси риб-бентофагів – 7,3 кг/га.
Вища водна рослинність на водоймі розвинута в
значній мірі і представлена як прибережним поясом повітряно-водної рослинності
(очерет, рогіз), так і м'якою зануреною рослинністю (рдесники пронизанолистний,
блискучий, гребінчастий). Занурена рослинність утворює щільні поля. Ступінь
заростання може бути оцінений як 80% площі водного дзеркала.
Первинний склад іхтіофауни Каховського
водосховища сформувався за рахунок видів, що перебували в р. Дніпро в зоні
затоплення із істотним впливом напівпрохідних форм Нижнього Дніпра та
Дніпровсько-Бузької естуарної системи. До складу іхтіофауни в перші роки
існування водосховища входило 47 видів, основними з яких були плоскирка,
синець, лящ, чехоня, тюлька, верховодка, окунь. В останні 10 років в складі
іхтіофауни Каховського водосховища зафіксовано 31 види риб, які відносяться до
10 родин (оселедцеві, щукові, коропові, в’юнові, сомові, колючкові, іглицеві,
атеринові, окуневі, бичкові), з яких промислове значення мають біля 20 видів.
Одним з основних чинників антропогенного впливу
на іхтіофауну Каховського водосховища є організація рибовидобувного промислу.
Максимальна промислова рибопродуктивність
водосховища – 46,5 кг/га – була досягнута у 1989 р. (в основному за
рахунок плітки, тюльки та товстолобів), надалі улови аборигенних видів риб
різко знизились и на початок поточного сторіччя рибопродуктивність становила
всього 9,4 кг/га. При цьому відбулась зміна видового складу промислових уловів.
Якщо у період 1991–1995 рр. питома вага заходів з штучного відтворення іхтіофауни
у формуванні промислової рибопродукції досягала 50%, то в останні десять років
основу ресурсної бази промислу складають аборигенні види.
Загальний промисловий вилов риби на Каховському
водосховищі, який за період 2001–2005 рр. мав тенденцію до збільшення: з 1,7
тис. т до 2,4 тис. т, у 2006 р. дещо зменшився – до 2136 т. В основному це
відбулось за рахунок зниження улову тюльки, тоді як вилов основних частикових
видів залишається стабільним, або зростає.
Сформована на сьогодні ресурсна база промислу Каховського
водосховища в певній мірі не відповідає структурі промислових уловів. Як
зазначалось вище, це в значній мірі пов'язане з організацією промислу та
кон’юнктурою ринку збуту виловленої риби.
Основним фактором позитивного впливу на зростання
валового вилову, було збільшення чисельності та іхтіомаси сріблястого карася,
негативного – зменшення технічної інтенсивності лову тюльки та скорочення
запасів плітки.
Вилов інших основних промислових видів протягом
останніх 10 років характеризується значною стабільністю. Рибопродуктивність
водосховища в 2008 р. становила 11,3 кг/га, що відповідає рівню середньої
по каскаду.
2. Матеріали та методи досліджень
2.1 Предмет та матеріали досліджень
Предмет дослідження – розмірно-віковий та статевий склад популяції
ляща Каховського водосховища.
Об’єкт дослідження – лящ Каховського водосховища.
Дипломна робота складається із матеріалів власних досліджень,
матеріалів Головного державного управління охорони, використання і відтворення
водних живих ресурсів та регулювання рибальства в Запорізькій області.
Власні дослідження проводилися протягом весняного періоду 2006 та
2009 років на іхтіологічних КСП (с. Малаєкатеринівка Запорізького району
Запорізької області).
В ході виконання досліджень було проаналізовано: із промислових
уловів – 2,4 тис. екземплярів плітки, із уловів контрольних сіток – 4,988 тис.
екземплярів. Проведено повний біологічний аналіз 200 екземплярів. Довжина риби
визначалася із точністю до 0,5 см, маса – до 1 г.
Огляд наукової літератури здійснювався з використанням фондів
провідних бібліотек Запорізької області та України: наукової бібліотеки
Запорізького національного Університету, Запорізької обласної бібліотеки ім.
Горького, Національної бібліотеки ім. Вернадського (м. Київ), наукової
бібліотеки Інституту рибного господарства Української аграрної академії наук.
2.2 Методи дослідження
На
контрольно-спостережних пунктах збір проб здійснювався стандартним набором
ставних сіток з кроком вічка а=30, 36, 40, 50, 60, 70, 75, 80, 90, 100, 110,
120 міліметрів, ставними неводами з кроком вічка 40 мм.
З уловів контрольних сіток оброблялися усі особини ляща. Для
повного біологічного аналізу відбиралися по 10 екземплярів риб кожного
розмірного класу. Розмірний клас становив 1 см. Розмірний ряд особин ляща
складався із мінімального та максимального розміру, які було виявлено в
знаряддях лову протягом періоду дослідження.
Проби для визначення плодючості відбиралися у самок із IV стадією
зрілості статевих продуктів в весняний донерестовий період при скупченнях
плідників на місцях нересту. Із передньої, середньої та задньої частини ястика
відбиралася частка ікри, змішувалась та відважувався 1 грам. Фіксування велося
за стандартними методиками [27, 28].
Підрахування ікри велося під бінокуляром МБС-1 з вимірюванням
діаметру ікринки (збільшення в 12 разів).
Визначення видового складу молоді проводилося у відповідності до
загальноприйнятих методик [29].
Визначення
морфологічних відмінностей між самцями та самками велося за загально прийнятими
методиками [30, 31, 32, 33, 34, 35,].
Для визначення віку риб використовувались стандартні методики:
підрахування вікових кілець на лусковій пластинці [36, 37].
З метою
визначення стану любительського рибальства дослідження проводили відповідно до «Методичних
вказівок з вивчення впливу любительського рибальства на стан рибних запасів
внутрішніх водойм», розробленими Науково-дослідним інститутом озерного і
річкового господарства модифікованих нами з урахуванням особливостей Каховського
водосховища та сучасних технічних можливостей [38]. Для визначення видів риб
використовувалися загальновідомі визначники [32, 33, 34, 35, 39, 40].
Для визначення
кількості рибалок, обсягу і якісного складу уловів любителів використовувалось
анкетне опитування та безпосередній облік рибалок і їхніх уловів на водосховищі.
Облік риболовів любителів на водоймі здійснювався
в будні та вихідні дні не менш ніж один раз на сезон.
Одночасно з обліком здійснювався збір інформації
з якісного та кількісного складу уловів з заповненням індивідуальної риболовної
картки.
Для одержання статистично достовірних даних
проводився індивідуальний облік уловів в день спостережень не менш ніж у 25
рибалок.
Кількість риби визначалась за формулою:
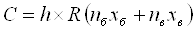 (2.1)
(2.1)
де С – кількість риби, що виловлюється,
(кг.);
h – середня тривалість завершеної риболовлі
(година.);
R – середня інтенсивність лову даного виду (шт./людино-година)
nб – Середнє число рибалок
на водосховища (його ділянці) у будній день;
xб – число будніх днів у
місяці;
nв – те ж для вихідного
дня;
xв – число вихідних днів у
місяці.
Середня інтенсивність лову визначається щомісяця
для кожного виду риб виходячи з результатів інтерв'ю й анкетного опитування по
формулі:
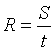 (2.2)
(2.2)
де R – інтенсивність лову;
S – загальна кількість риби, виловленої всіма
опитаними рибалками-аматорами згідно рибальських карток (кг/шт.);
t – загальна кількість годин, витрачених цими
рибалками на вилов усієї кількості риби (година).
Тривалість завершеної риболовлі підраховується на
підставі даних анкетного опитування любителів.
Для визначення риболовного навантаження на
водойму використовувалась формула:
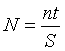 (2.3)
(2.3)
де N – риболовне навантаження (година /га
рік);
п – кількість проведених риболовель;
t – середній час завершеної риболовлі;
S – площа водосховища, його ділянки.
Верстання та редакція тексту здійснювалась у
текстовому редакторі Microsoft Word 2007.
2.3 Статистична обробка
результатів
Статистичну обробку
інформації проводили за загальноприйнятими методиками [55].
Помилка репрезентативності
середнього значення розраховувалась за формулою 2.4:
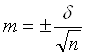 (2.4)
(2.4)
де m – помилка
репрезентативності;
δ – середньоквадратичне
відхилення.
Середньоквадратичне
відхилення визначали за формулою 2.5:
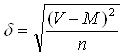 (2.5)
(2.5)
де V – абсолютне значення признаку (см);
М – серед неарифметичне значення ряду вибірки.
3. Експериментальна частина
3.1
Розмірний склад популяції ляща Каховського водосховища
Протягом 2004 та 2005
років в популяції ляща в верхній частині Каховського водосховища переважали
особини довжиною 34–35 см, їх частка в контрольних уловах, що проводились
на весні становила 28% (таблиця 3.1, рисунок 3.1).
Найближчі розмірні
групи 36–37 см склали 6,9% в 2004 році та 15,56% – в 2005, а 32–33 –
близько 18,77% та 10,46% відповідно. Середній розмір протягом 2004–2005 років
має стабільну динаміку та складає близько 37 см.
В 2006 році
спостерігається дещо інша ситуація. В контрольних уловах більшість складають
особини 32–33 см (промислова міра на ляща встановлена на рівні
32 см). Частка особини довжиною 34–35 см склала 19%.
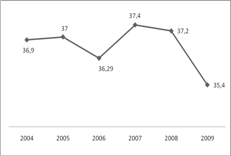
Рік
Рисунок 3.1 – Динаміка
середнього розміру ляща в контрольних уловах на Каховському водосховищі у 2004–2009
роках
Проте в уловах з 2006
року зустрічаються особини крайніх розмірних класів – 52–57 см. Середній
розмір знизився до 36 см.
Протягом 2007 року
середній розмір ляща в контрольних уловах збільшився майже на 1 см.
Більшість складали особини розміром 36–37 см. В останні два роки відсоток
особин цього розмірного класу стабільно знижується.
У 2008 році більшість
також припадає на особин розміром 36–37 см., проте у 2007–2008 роках
значно зменшилась частка особин з промисловим розміром до 32 см. Середній
розмір у 2009 році знизився до 35,4 см.
3.2
Віковий склад популяції ляща Каховського водосховища
В середньому за весь
період 2004–2009 роки найбільший відсотковий склад становили особини 6–8 років.
Так, с особини 6 – річного віку становили, в середньому – 25,5%, особини 5 –
річного віку – становили 22,1%, а особини 6 – років – 22,8%. Також
спостерігалася значна кількість особин ляща 8 – років – 13,1%. (Таблиця 3.2,
рисунок 3.2).
Протягом 2004 року в
популяції ляща Каховського водосховища переважали особини 7-річного віку (24,0%)
(рис. 3.2). на другому місці за відсотковим складом знаходились особини 6
– річного віку (23,56%), а 4 – річні особини складали лише 1,19%. інші вікові
групи – 9 та 10-річні особини становили, відповідно – 9,2% та 8,68%. лише по
1,06% та 0,77% були представлені 11 – та 12 – річні особини ляща. Середній вік
склав 6,99 років.
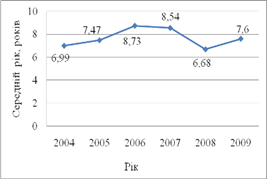
Рисунок 3.2 – Динаміка
середнього розміру лящ в контрольних уловах на Каховському водосховищі у 2004–2009
роках
У 2005 році віковий склад популяції ляща дещо
змінюється. Всього нараховувалось 9 вікових категорій – з 4 по 12. Так, 5 – річні
особини складають вже 4,56% від загальної кількості проаналізованих особин.
Найбільші показники за відсотковим складом становлять 7 – та 8-річні особини –
відповідно 27,19 та 38,0%. Значно збільшується кількість 7 та 8 річних особин
ляща – 8,3 та 25,87%, відповідно.
У 2006 році в контрольних уловах вже
зустрічаються особини 17 вікових категорій. Найбільший відсоток припадав на
особин 5-річного віку – 19,53%. Кількість 6 – річних особин знаходилася
практично на одному рівні і становила – 16,27%. Відсоток особин, що досягли 7
та 8 років значно зменшився. 3-річні особини становили 0,51%, 4 – річні особини
– 4,34%. Старші вікові групи, в порівнянні з минулими роками значно збільшилися
та становили: 11-річні особини – 4.34%, 12-річні особини – 6,65%, а 11 – річні
особини – 6,65%.
У 2007 році більшість складали особини, що
досягли 7 років – 20,22%. Кількість 8 – річних особин, зросла і становила 18,11%.
На третьому місті за віковим складом знаходилися 9 – річні особини – 16,9%.
Середній взважений показник склав 8,57%.
У 2009 році в контрольних уловах на Каховському
водосховищі нараховувалось 17 вікових категорій: з 3 по 19 років. Найбільша
частка припадала на особин, що досягли 5 років – 15,4%. На другому та третьому
місцях – особини з віком 4 та 7 років, на їх частку припадало 13,9% та 13,4%
відповідно. Середній зважений показник дещо збільшився у порівнянні з
аналогічним показником 2008 року та склав 7,6 років.
Індивідуальні
біологічні показники ляща (середня довжина та маса по вікових групах,
вгодованість (1,8–2,2 за Фультоном) свідчать про нормальні умови нагулу, а
розподіл улову за кроком вічка – 65,6% загальної його кількості та 64,6% маси
забезпечувалось за рахунок сіток з а=75–80 мм – про можливість організації
ефективного облову найбільш оптимальними для даного виду знаряддями лову. У
2009 р. показники, які характеризують стан та експлуатацію запасів ляща Каховського
водосховища становили: К заг. см – 36,5; К прир. см –
14,6; Квилову – 21,9.
Таблиця 3.3 – Біологічні
показники ляща верхньої частини Каховського водосховища в контрольних уловах
ставних сіток з а=30–120 мм у 2009 р.
| Вікові групи |
Віковий склад, % |
Довжина, см |
Маса, г |
| 4 |
1,2 |
27,7 |
406 |
| 5 |
2,9 |
30,5 |
600 |
| 6 |
6,3 |
32,7 |
732 |
| 7 |
12,2 |
35,0 |
915 |
| 8 |
18,5 |
37,0 |
1057 |
| 9 |
16,3 |
38,8 |
1240 |
| 10 |
18,8 |
40,8 |
1392 |
| 11 |
12,9 |
42,1 |
1574 |
| 12 |
6,3 |
44,1 |
1709 |
| 13 |
1,7 |
45,3 |
1844 |
| 14 |
1,6 |
47,6 |
2314 |
| 15 |
1,2 |
49,0 |
2422 |
| Середньовиважені |
9,1 |
38,7 |
1249 |
| Кількість екз. |
423 |
|
|
Впродовж 2005–2009
років в уловах контрольних сіток на Каховському водосховищі спостерігається
незначне коливання середнього розміру особин ляща. Так у 2005 році він склав 37 см.,
у 2006 знизився майже до 36 см. протягом 2007 та 2008 років виловлювався
лящ відповідно 37, 4 та 37,2 см (таблиця 3.3).
3.3 Статевий склад популяції та плодючість ляща
на Каховському водосховищі
Абсолютна індивідуальна плодючість (ІАП) самиць
ляща в середньому склала 114,8 тис. ікринок, відносна плодючість (ВП) – 0,211
ікр./кг.
Таблиця 3.4 – Плодючість ляща Каховського
водосховища в 2009 році
| Довжина риб, см |
Середня маса риб, кг |
ІАП, шт.ікринок |
ВП, ікр./кг |
| 15 |
0,085 |
24700 |
288 |
| 16 |
0,100 |
19000 |
190 |
| 17 |
0,100 |
17566 |
176 |
| 18 |
0,137 |
38025 |
270 |
| 19 |
0,151 |
41568 |
289 |
| 20 |
0,185 |
29193 |
165 |
| 21 |
0,215 |
65780 |
310 |
| 22 |
0,241 |
61084 |
255 |
| 23 |
0,294 |
52164 |
185 |
| 24 |
0,326 |
50703 |
161 |
| 25 |
0,329 |
70589 |
209 |
| 26 |
0,430 |
74884 |
175 |
| 27 |
0,446 |
96005 |
212 |
| 28 |
0,481 |
101614 |
211 |
| 29 |
0,538 |
106800 |
194 |
| 30 |
0,630 |
158218 |
234 |
| 31 |
0,682 |
145407 |
212 |
| 32 |
0,776 |
163459 |
210 |
| 33 |
0,803 |
170760 |
199 |
| 34 |
0,989 |
255572 |
– |
| 35 |
1,112 |
250360 |
224 |
| 36 |
1,131 |
231844 |
203 |
| 37 |
1,250 |
222704 |
177 |
| 38 |
1,383 |
271660 |
196 |
| 39 |
1,440 |
288580 |
200 |
Абсолютна
індивідуальна плодючість плітки Каховського водосховища за даними досліджень в
2009 p. коливалась від 24,7 тис. ікринок у самиць довжиною 15 см і вагою
0,085 кг, до 288,6 тис. ікринок у самок довжиною 39 см і вагою 1,440 кг. Значення
індивідуальної абсолютної плодючості позитивно корелює з масою й довжиною тіла
риб – 0,98 і 0,95.
Проведеними
дослідженнями, зокрема за даними контрольних ловів у переднерестовий і
нерестовий період співвідношення самців і самиць у популяції ляща Каховського
водосховища за період 2008–2009 років у середньому було близьке до 1:1, проте у
верхній і середній частинах водоймища серед особин старших вікових груп (більше
6-ти років) співвідношення самців і самиць було зрушено у самиць з перевагою
останніх приблизно на 40% від загального числа досліджених особин.
4.
Охорона праці
Під час виконання дипломної роботи дослідження
проводилися в польових умовах.
В ході роботи використовувалися наступні засоби
індивідуального захисту: костюм х/б, костюм рибацький водостійкий, плащ
гумовий, фартух гумовий, костюм теплий, чоботи гумові, печатки латексні.
Безпосередньо перед початком робіт ми одягали індивідуальні
засоби захисту, ознайомлювалися з завданням керівника практики та прогнозом
погоди, проходили цільовий інструктаж з охорони праці.
Під час пересування берегом дотримувались
наступних правил безпеки:
-
заборонялося перестрибувати з камінню на камінь;
-
переходили небезпечні ділянки (струмки, тощо) виключно з
використанням страхування одного з членів групи, лише
після остаточного переходу небезпечної ділянки одним з членів групи відбувався
перехід інших;
-
в темний час доби здійснювали підсвічування шляху ліхтарем;
Під час виходу на воду на маломірних судах
дотримувались таких правил безпеки:
-
під час посадки та висадки дотримувались відповідної черги;
-
всі члени групи були одягнені в рятувальні нагрудники або жилети
та не знімали їх до виходу з судна;
-
на судні заборонялося стояти, сідати на борти та ранцеву дошку;
-
пересуватися та мінятися місцями без дозволу старшого на судні;
-
палити та користуватися відкритим вогнем.
Після закінчення роботи здійснювали чищення
засобів індивідуального
захисту та спецодягу, після чого складували у
відведене для них місце.
Після контакту з рибою руки ретельно промивалися
теплою водою з милом.
Добір загиблої риби і мулу в місцях загибелі
здійснювали в целофанові мішки, що зав'язували, чи в іншу тару, що
закривається.
Проби води відбираються в герметичну тару.
Забороняється брати загиблу рибу голими руками.
Під час приймання зарибку на рибоводних
підприємствах дотримувалися правил з охорони праці, що діють на даному
підприємстві.
При роботі на комп'ютерах, до складу яких входять
відеодисплейні термінали (ВДТ), на працівників можуть діяти наступні небезпечні
і шкідливі виробничі чинники: фізичні (підвищений рівень шуму на робочому місці
(від вентиляторів процесорів і ау-діоплат);
підвищене значення напруги в електричному ланцюзі|цепі|,
замикання якої може відбутися через тіло людини; підвищений рівень статичної
електрики; підвищений рівень електромагнітного випромінювання; підвищена
напруженість електричного поля; пряма і відображена від екранів мерехтіння,
несприятливий розподіл яскравості в полі зору); психофізіологічні (фізичні
перевантаження статичної і динамічної дії; нервово-психічні перевантаження
(розумове перенапруження, перенапруження аналізаторів, монотонність праці,
емоційні перевантаження).
Основним атребутомобладнанням
робочого місця користувача комп'ютера є: ВДТ, клавіатура, робочий стіл, стілець
(крісло); допоміжним – пюпітр, підставка для ніг, шафи, полиці і ін.
Робочі місця з ВДТ
розташовані на відстані не менше 1,5 м від стіни з вікнами, від інших стін
– на відстані 1 м, між собою – на відстані не менше 1,5 м. Щодо вікон
робоче місце розміщували так, щоб природне світло падало на нього збоку,
переважно зліва та так, щоб уникнути
попадання в очі прямого світла. Джерела освітлення розташовували з обох сторін
екрану паралельно напряму погляду.
Щоб уникнути світлових
відблисків від екрану, клавіатури у напрямі очей користувача, від світильників
загального освітлення або сонячних променів ми застосовували сітки,
антивідблисків, спеціальні фільтри для екранів, захисні козирки, на окнах –
жалюзі.
При роботі з текстовою
інформацією (у режимі введення даних, редагування тексту і читання з екрану
ВДТ) найбільш фізіологічним є зображення чорних знаків на світлому (білому)
фоні.
Ми розміщували ВДТ на
робочому місці так, щоб поверхня екрану знаходилася в центрі поля зору на
відстані 400–700 мм від очей користувача. Рекомендується розміщувати
елементи робочого місця так, щоб підтримувалася однакова відстань від очей
користувача до екрану, клавіатури, пюпітра.
Робота комп'ютерів
супроводжується електромагнітним випромінюванням низьких рівнів, інтенсивність
якого зменшується пропорційно квадрату відстанівід екрану. Оптимальним, для
того, що працює, є відстань 500 мм від екрану ВДТ.
Зручна робоча поза при
роботі на комп'ютері забезпечується регулюванням висоти робочого столу, крісла
і підставки для ніг.
Раціональною робочою
позою може вважатися таке положення тіла, при якому ступні працівника
розташовані горизонтально на підлозі або на підставці для ніг, стегна
зорієнтовані в горизонтальній площині верхні частини рук – вертикально, кут
ліктьового суглоба коливається в межах 70–90о,
зап'ястя зігнуті під кутом не більше 200,
нахил голови – в межах 15–200.
Для нейтралізації зарядів
статичної електрики в приміщенні де виконуються роботи на комп'ютерах, зокрема
на лазерних і светодіодних принтерах, ми
збільшували вологість повітря за допомогою кімнатних зволожувачів. Не
рекомендується носити одяг з синтетичних матеріалів.
Під час роботи за
комп’ютером клавіатуру розміщували стійко на робочому столі, не допускаючи її
хитання. Разом з тим повинна була передбачена можливість її поворотів і переміщень.
Положення клавіатури і кут її нахилу відповідали побажанням користувача.
Якщо в конструкції
клавіатури не передбачено простір для опори долонь, то її слід розміщувати на
відстані не менше 100 мм від краю столу в оптимальній зоні моторного поля.
При роботі на клавіатурі сиділи прямо, не напружуючись.
Тривалість безперервної
роботи за ВДТ без регламентованої перерви не перевищувала 2 годин.
Закінчуючи роботу ми
записували файл в пам'ять комп'ютера. Виходили з програмної оболонки. Вимикали
принтер, інші периферійні пристрої, ВДТ і процесор, стабілізатор, якщо
комп'ютер підключений до мережі через нього. Штепсельні вилки виймали з
розеток. Накривали клавіатуру кришкою для уникнення попадання на неї пилу.
Приводили в порядок робоче місце. Оригінали і інші документи клали в ящик
столу. Ретельно мили руки теплою водою з милом. Виключали кондиціонер
освітлення і загальне електроживлення підрозділу.
Людині, що тонула
надається наступна перша допомога:
З постраждалого необхідно бистро зняти мокрий
одя, очистити йому рот від грязі. Видалити з дихальних шляхів та шлунка воду.
Для цього треба стати на одне коліно, покласти постраждалого поперек другого
свого коліна та натискати руками на спину. Після цього провести штучне дихання
та непрямий массаж серця. Сенс непрямого масажу серця є в ритмічному
натискуванні між грудною клітиною та хребтом. При проведенні непрямого масажу
серця постраждалого кладуть спиною на рівну тверду поверхню. Особа, яка надає
допомогу, стає збоку, нащупує нижній край грудини і на 2–3 пальці вище кладе на
неї опорну частину долоні, зверху прикладає іншу долонь під прямим кутом до
першої, при цьому пальці не повинні торкатися грудної клітини. Енергійними
ритмічними рухами натискують на грудну клітину з такою силою, щоб прогнути її в
сторону хребта на 4–5 см. Частота натискувань – 60–80 раз./ хв.
Ефективність непрямого масажу серця оцінують по з'явленню пульсації на сонних
та променевих артеріях, підвищення артеріального тиску, звуженню зіниць та
з'явленню реакції на світло, зникненню блідості.
Якщо хворий почав дихати укласти його в ліжко,
тепло вкрити та дати йому теплого чаю або кави. Одночасно з наданням першої
допомоги треба викликати лікаря. Транспортування постраждалого з зупинкою
дихання та серцевих скорочень може бути проведене лише після відновлення
серцевої діяльності та дихання, або в спеціалізованій машині швидкої допомоги.
При забиванні необхідно покласти постраждалого, а
на забите місце накласти пов'язку та поверх неї покласти холодний компрес.
При порізах – очистити рану марлевим тампоном та
змастити йодом або накласти пов'язку. Не дозволяється рану промивати водою та
зав'язувати хусткою, які вже використовувалися.
При пораненні руки насамперед необхідно здійняти
її в гору, а при пораненні ноги лягти та також здійняти ногу до гори. Потім
накласти вище рани жгут з реміня або полотенця та утримувати поки не зупиниться
кровотеча.
Забороняється накладати жгут постраждалому більше
ніж на дві години. При пораненнях та переломах кісток черепу, кінці рани
змазати йодом, на рану накласти стерильну пов'язку. Після надання першої
допомоги доставити постраждалого в лікарню.
Відмороження – ушкодження тканин в результаті дії
низької температури. Перша допомога у разі відмороження полягає в негайному
зігріванні постраждалого, для чого його необхідно як можна швидше перевести в
тепле приміщення. Насамперед треба зігріти відморожену частину тіла, відновити
в ній кровообіг. Найбільшого ефекту й безпеки можна досягти за допомогою
теплових ванн. За 20–30 хвилин температуру води поступово збільшують від 20 до
40 °С, при цьому кінцівки ретельно відмивають від забруднень. Після ванни
ушкодженні частини осушити, закрити стерильної пов'язкою та тепло вкрити.
Відморожені частини тіла забороняється розтирати
снігом, змащувати жиром, або мазями. Велике значення при наданні першої
допомоги мають заходи з загального зігрівання постраждалого. Йому дають гарячий
чай, каву, молоко.
Постраждалого необхідно якомога швидше доставити
до лікарні. При транспортуванні слід прийняти всі заходи щодо попередження
повторного охолодження [41].
Висновки
1.
Спостерігаються зміни у розмірному складі ляща в контрольних
уловах на Каховському водосховищі впродовж 2005–2009 рр. більшість припадає на
особин з промисловим розміром 36–37 см., проте у 2007–2009 роках значно зменшилась
частка особин з промисловим розміром до 32 см.
2.
В середньому за весь період 2004–2009 років найбільший відсотковий
склад становили особини 6–8 років. Так, с особини 6 – річного віку становили, в
середньому – 25,5%, особини 5 – річного віку – становили 22,1%, а особини 6 –
років – 22,8%. Також спостерігалася значна кількість особин ляща 8 – років –
13,1%.
3.
Співвідношення самців і самиць в популяції ляща Каховського
водосховища за період 2008–2009 років у середньому було близьке до 1:1, однак у
верхній і середній частинах водоймища серед особин старших віків (більше 6-ти
років) співвідношення самців і самиць було зрушено у самиць з перевагою
приблизно на 40% від загального числа досліджених особин.